Trochę trudniejsze zagadnienie ale spróbuję sobie jakoś poradzić z jak najprostszym przedstawieniem tematu. Nie mogę znaleźć polskiego odpowiednika do określenia cross-over więc pozostanę przy orginalnym nazewnictwie.
Cross-over odgrywa przede wszystkim ważną rolę przy łączeniu mutacji umiejscowionych na tym samym chromosomie. To jest coś, co najczęściej zauważamy w mutacjach związanych z płcią. Mówiąc prościej, możemy stwierdzić, że podczas cross-over fragmenty DNA (geny) są wymieniane z jednego chromosomu na drugi. Frans Alfons Janssens (1863 - 1924), profesor biologii i cytologii na Katolickim Uniwersytecie w Leuven, odkrył w 1909 roku, że podczas generowania komórek rozrodczych chromatydy w pewnym momencie zachodzą na siebie. Nazwał miejsce, w którym to występuje chiasmą. Uważał to za ważną przyczynę istnienia jakichkolwiek mutacji i wymiany genów.
Cross-over w działaniu
Pasujące do siebie chromosomy wyszukują się nawzajem podczas profazy i mejozy (będzie o tym w innym wpisie). W tym momencie para chromosomów składa się z czterech siostrzanych chromatyd (tetrad). Te chromatydy zostają „zgniatane” podczas synapsy. Z tego powodu w pewnym momencie się zrywają ("łamią"). Przejście następuje, gdy te uszkodzone fragmenty DNA przyczepiają się do chromatydy obok, w ten sposób mutacje jednego chromosomu są przekazywane do drugiego. Tę rekombinację genów po raz pierwszy zademonstrował angielski badacz Bateson (1861-1921). Ponieważ w tym momencie są cztery chromatydy razem w tetradzie i są różne lokalizacje krzyżowe lub chiasma, istnieją oczywiście różne możliwości tego, jak to pęknięcie lub rekombinacja może wystąpić.
Wizualna reprezentacja cross-over
Aby wyjaśnić to nieco prościej, oto przedstawienie dość prostego cross-over na rysunku, ale proszę pamiętać, że możliwe jest więcej niż jedno cross-over i że jest to tylko losowy przykład.

Na rysunku wyraźnie widać, że w pobliżu chiasma chromatydy pękają i ponownie przyczepiają się do drugiego chromosomu. Zgodnie z umowami międzynarodowymi nie tworzy się nowych nazw dla kombinacji mutacji. Więc również nie dla fenotypów spowodowanych cross-over. Kombinacje są wskazane przez umieszczenie łącznika między nazwami zaangażowanych mutacji. Na przykład kombinacje cynamonu i SL ino będą cynamonem-SL ino. Opaline i cynamon zamienią się w opaline-cynamon itd. Znowu zasada jest taka, że najmniej „zredukowana mutacja” jest wymieniana jako pierwsza. Tak więc w cynamonie czarny pigment staje się tylko brązowy i jest bardziej obecny niż w SL ino green, więc cynamon jest wymieniony jako pierwszy w nazwie. Opaline tak naprawdę nie ma redukcji, ale rearanżację pigmentu, więc w tym połączeniu będzie to przed cynamonem itp.
Cross-over w praktyce
Wyraźnym praktycznym przykładem cross-over jest opaline-SL ino u Agapornis roseicollis (czasami niestety nadal nazywany rubino). Jest to kolejne cross-over dwóch recesywnych mutacji związanych z płcią. Aby lepiej to zilustrować, ponownie użyjemy rysunku.
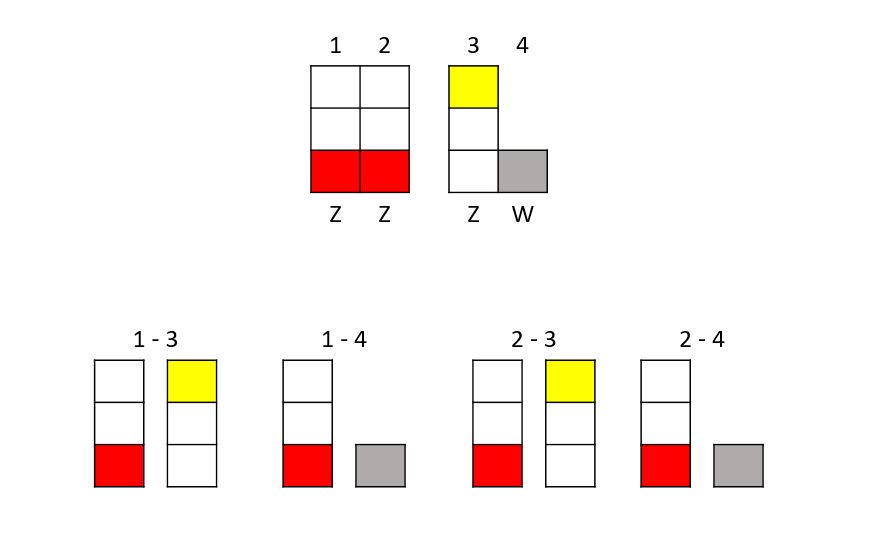
Mamy opalowego samca (bloczki czerwone) i samicę SL ino zieloną (bloczki żółte).
Kiedy połączymy te ptaki w pary, otrzymamy następujące kombinacje:
Znowu mamy cztery możliwe wyniki:
1-3: Młode mają dwa chromosomy Z. Jeden ze zmutowanym genem opaline i jeden ze zmutowanym genem SL ino. Stąd ptak jest samcem, zielonym, podwójnym szpaltem, więc zielony/SL ino/opaline.
1-4: Ta kombinacja daje chromosom Z i W, a więc samicę. Obecny jest tylko zmutowany gen opaline, więc jest ona opaline-zielona.
2-3: Znowu dwa chromosomy Z, a więc samiec. Podobnie jak w pierwszej kombinacji, jeden chromosom będzie miał zmutowany gen opaline, a drugi zmutowany gen SL ino. Tak więc ptak będzie zielony podwójny szpalt (zielony/SL ino/opaline).
2-4: Podobnie jak w przypadku drugiej kombinacji, będzie to samica opaline.
Samce z tego skojarzenia są podwójnymi szpaltami. Samica nigdy nie może zostać podwójnym szpaltem, ponieważ ma tylko jeden aktywny chromosom Z w mutacji sprzężonej z płcią.
Możemy pójść o krok dalej i połączyć podwójnego szpalta samca z samicą typu dzikiego.

To da nam następujące wyniki TEORETYCZNE:
1-3: 25% prawdopodobieństwo samca, szpalta zielony/opaline
1-4: 25% prawdopodobieństwo samica zielona opaline
2-3: 25% prawdopodobieństwo samiec szpalt zielony/SL ino
2-4: 25% prawdopodobieństwo samica SL ino zielona
To wszystko może wydawać się logiczne, ale widzimy, że w rzeczywistości wyniki hodowlane nieco się różnią i regularnie rodzą się młode samiczki opaline-SL ino. Teoretycznie nie jest to możliwe, ale w rzeczywistości są to możliwe wyniki:
17,5% prawdopodobieństwo zielony/SL ino (samce)
17,5% prawdopodobieństwo zielony/opaline (samce)
7,5% prawdopodobieństwa zielonych (czystych) samców
7,5% prawdopodobieństwa zielonych/opaline-SL ino (samców)
17,5% prawdopodobieństwo SL ino zielony
17,5% prawdopodobieństwa samic opaline
7,5% prawdopodobieństwa zielony (czysty)
7,5% prawdopodobieństwo wystąpienia samic opaline-SL ino
Jak to jest możliwe?
Kiedy samiec jest podwójnym szpaltem, może się zdarzyć, że podczas mejozy (kiedy powstają komórki rozrodcze) następuje cross-over, w którym obie mutacje będą zlokalizowane na tym samym chromosomie.
Aby to wyjaśnić, użyjmy rysunków:
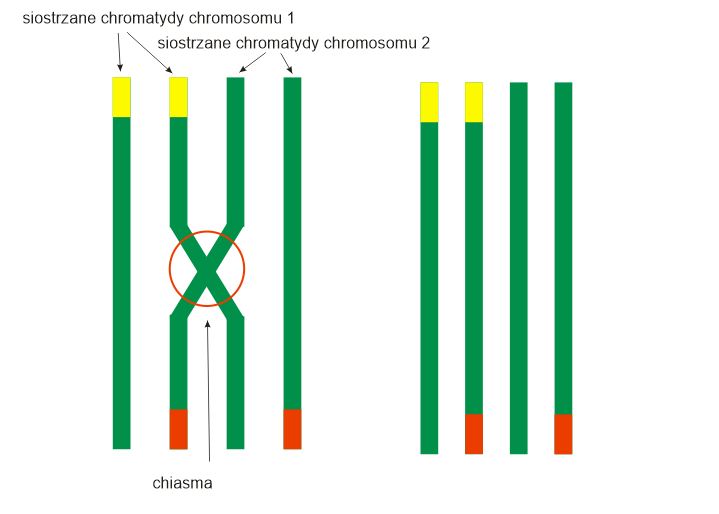
Na tym rysunku widzimy cztery chromatydy podwójnego szpalta samca (zielony/SL ino/opaline). Dwie po lewej są nosicielami żółtego bloku reprezentującego gen SL ino. Dwie po prawej to nośniki opaline reprezentowane przez czerwone bloki.
Jak widać na rysunku, dwa zmodyfikowane chromatydy powstają podczas tworzenia komórek rozrodczych z powodu tego cross-over.
Jedna chromatyda jest nosicielem dwóch mutacji (opaline i SL ino), podczas gdy inna nie ma już żadnych mutacji. Jeśli ta zmodyfikowana chromatyda trafi do młodego samca, będzie on zielony/opailne-SL ino (czynniki są ze sobą powiązane). Jeśli trafi na młodą samiczkę będzie ona opaline-SL ino.
Gdy nastąpiło cross-over, moglibyśmy, świadomie lub nie, sparować takiego samca szpalta z samicą typu dzikiego i (teoretycznie) moglibyśmy mieć następujące kombinacje:
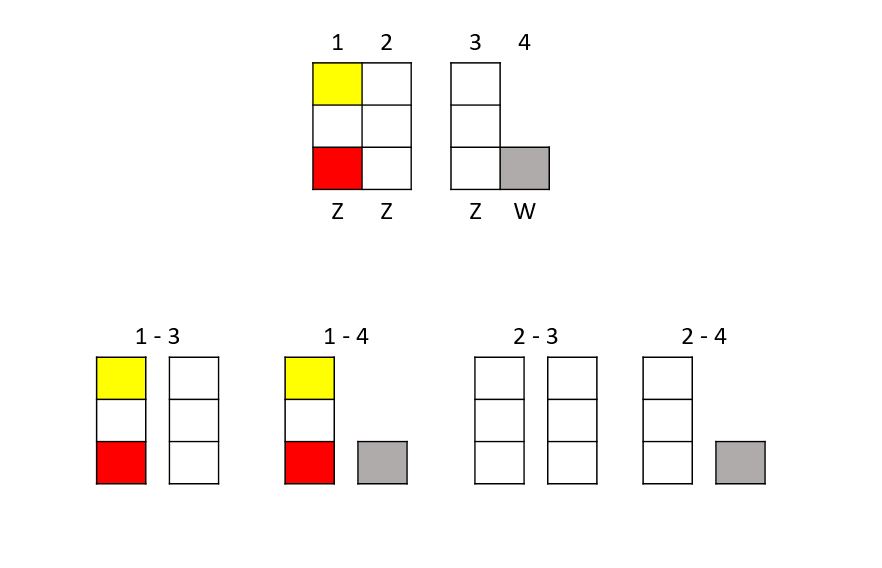
1-3: 25% prawdopodobieństwo zielonego samca, szpalta na powiązane czynniki opaline-SL ino
1-4: 25% prawdopodobieństwo samicy opaline-SL ino
2-3: 25% prawdopodobieństwo zielonego samca
2-4: 25% prawdopodobieństwo zielonej samicy
Ale te wyniki również mogą się różnić.
Gdy nastąpi cross-over, które łączy dwa czynniki, te połączone czynniki można po prostu oddzielić, ponownie przez cross-over, tworząc oddzielne mutacje.
W rzeczywistości połączenie skutkuje:
7,5% prawdopodobieństwo zielonych/SL ino samców
7,5% prawdopodobieństwo zielonych/opaline samców
17,5% prawdopodobieństwo zielonych samców
17,5% prawdopodobieństwo samców zielonych/opaline-SL ino (czynniki powiązane)
7,5% prawdopodobieństwo SL ino zielonych samic
7,5% prawdopodobieństwo opaline zielonych samic
17,5% prawdopodobieństwa opaline-SL samic (czynniki powiązane)
17,5% prawdopodobieństwo zielonych samic
Więc ci, którzy myślą, że można poszufladkować naturę w małych pudełkach, powinni pomyśleć dwa razy.
Ważne
Ponieważ przykłady używane do cross-over są zwykle mutacjami SL (sprzężonymi z płcią), jest powszechnym nieporozumieniem, że cross-over występuje tylko w przypadku mutacji sprzężonych z płcią. To nie jest prawda. Wszystkie pary chromosomów podlegają cross-over. Problem polega na tym, że nie zawsze wiemy w chromosomach autosomalnych, kiedy na tym samym chromosomie występują dwie mutacje. Podręcznikowym przykładem jest dark factor. Wiemy na pewno, że znajduje się on na tym samym chromosomie co mutacja niebieska i dlatego konieczne jest cross-over, aby uzyskać zarówno niebieski, jak i dark factor na tym samym chromosomie.
Prawdopodobieństwo cross-over
Szanse, że nastąpi cross-over, zależą od odległości między obiema mutacjami. Im większa odległość między obiema mutacjami na chromosomie, tym większe prawdopodobieństwo, że chromosom pęknie gdzieś pośrodku i oczywiście nastąpi cross-over.
Odległość na chromosomie między różnymi mutacjami jest wyrażona w centymorganach. Termin ten nosi imię Amerykanina Thomasa Hunta Morgana (1866-1945), który na podstawie odkrycia Fransa Alfonsa Janssensa odkrył, w jaki sposób można obliczyć odległość między genami na chromosomach za pomocą częstotliwości wystąpienia cross-over mutacji.
U ptaków znana jest liczba częstości krzyżowania się mutacji. Oto przegląd znanych procentów cross-over u nierozłączek.
Na początek mutacje na chromosomie Z (mutacje sprzężone z płcią):
Cynamon i SL ino: 3%
Cynamon i opaline: 33%
SL ino i opaline: 30%
Na chromosomie autosomalnym:
Dark factor i niebieski: 7%
U nierozłączek cynamon i opaline są najbardziej oddalone od siebie na chromosomie Z. Mają one 33% występowania cross over. Najkrótsza odległość znajduje się między cynamonem a SL ino. U papug są one oddalone od siebie o zaledwie 3 centymorgany. Możliwość przejścia to zaledwie 3%.
Ważne jest, aby wiedzieć, że procent cross-over u nierozłączek jest inny niż u np. kanarków. Dzieje się tak, ponieważ chromosomy u kanarków ewoluowały z czasem w nieco inny sposób. Rezultatem jest inna sekwencja zasad DNA.
Cross-over nie jest przypadkiem!
Ludzie kiedyś myśleli, że cross-over były raczej wyjątkiem, ale teraz wiemy, że jest to naturalny sposób na urozmaicenie naszych genów. U ludzi występuje średnio od 50 do 60 przejść na mejozę. To średnio od 2 do 3 na chromosom. Jest to unikalny system utrzymania naszej różnorodności genetycznej. W rezultacie DNA każdej osoby jest wyjątkowe.


